Research Interests
Motivated by the challenges and opportunities created by global change, my research: (1) quantifies and develops mechanistic understandings of the distribution, structure, and dynamics of populations and communities over space and time; and (2) documents and understands how wildlife species and ecosystems respond to global change.
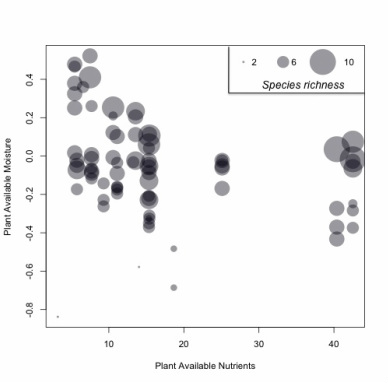 Large herbivore species richness in India.
Large herbivore species richness in India.
Species richness in space and time
I am interested in questions related to how species are distributed in space and time. I have investigated to what extent environmental variables influence the distribution of the species richness (number of species) of large mammalian herbivores in India. I analyzed the distribution and richness of the 16 large (>10 kg) herbivore species found in 76 protected areas (>200 km2) in sub-Himalayan India with respect to PAM (log10(rainfall/potential evapotranspiration)), PAN (soil cation exchange capacity), elevation, tree cover, and fire frequency. Using regression and null models, I found positive relations between PAM and the occurrences of the larger-bodied species (elephant and gaur), and negative relations between PAM and the occurrences of smaller-bodied species (chinkara, four-horned antelope and blackbuck). I also found positive relations between the occurrence of the smaller-bodied species and PAN. Large herbivore species richness in India is highest in Kanha and Indravati, areas with high PAN and intermediate PAM (Fig. on left). I found elevation, tree cover and fire frequency to be insignificant predictors of herbivore species richness, although elevation and tree cover explained the distribution of a few species. These results suggest that moisture and soil nutrients are important determinants of large herbivore species distribution and richness in sub-Himalayan India.
I am interested in questions related to how species are distributed in space and time. I have investigated to what extent environmental variables influence the distribution of the species richness (number of species) of large mammalian herbivores in India. I analyzed the distribution and richness of the 16 large (>10 kg) herbivore species found in 76 protected areas (>200 km2) in sub-Himalayan India with respect to PAM (log10(rainfall/potential evapotranspiration)), PAN (soil cation exchange capacity), elevation, tree cover, and fire frequency. Using regression and null models, I found positive relations between PAM and the occurrences of the larger-bodied species (elephant and gaur), and negative relations between PAM and the occurrences of smaller-bodied species (chinkara, four-horned antelope and blackbuck). I also found positive relations between the occurrence of the smaller-bodied species and PAN. Large herbivore species richness in India is highest in Kanha and Indravati, areas with high PAN and intermediate PAM (Fig. on left). I found elevation, tree cover and fire frequency to be insignificant predictors of herbivore species richness, although elevation and tree cover explained the distribution of a few species. These results suggest that moisture and soil nutrients are important determinants of large herbivore species distribution and richness in sub-Himalayan India.
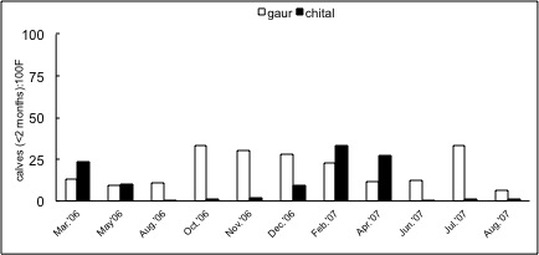 Mudumalai & Bandipur National Parks, South India, Mar '06 – Aug '07
Mudumalai & Bandipur National Parks, South India, Mar '06 – Aug '07
Bottom-up control of animal behavior: seasonal plant quality, body mass and the timing of parturition
Seasonal variation in forage availability and quality is understood to affect the annual timing of parturition in large herbivores. In India--where seasonal monsoonal rains define variation in forage availability and quality--chital Axis axis exhibit stronger seasonality in parturition than the larger gaur Bos taurus (Fig. on left). I hypothesized that this difference could be explained by forage requirements determined by body mass. Results showed that the minimum forage nitrogen content required by lactating gaur was available throughout the year, but that required by lactating chital was available for less than 40% of the year, i.e. during the early wet season, which coincides with the annual peak period of chital births. The three to four month spread of chital births, which begins in the dry season, implies that the period of highest quality is also important for females to replenish maternal reserves for future reproduction and help maximize the growth rate of neonates. This spread also indicates low synchrony of chital births and suggests that predator swamping was less important in influencing their timing of parturition. Insensitivity of the results to variable rainfall patterns lent support to the hypothesis that forage requirements determined by body mass might explain the variation in parturition strategies among large herbivores species.
Seasonal variation in forage availability and quality is understood to affect the annual timing of parturition in large herbivores. In India--where seasonal monsoonal rains define variation in forage availability and quality--chital Axis axis exhibit stronger seasonality in parturition than the larger gaur Bos taurus (Fig. on left). I hypothesized that this difference could be explained by forage requirements determined by body mass. Results showed that the minimum forage nitrogen content required by lactating gaur was available throughout the year, but that required by lactating chital was available for less than 40% of the year, i.e. during the early wet season, which coincides with the annual peak period of chital births. The three to four month spread of chital births, which begins in the dry season, implies that the period of highest quality is also important for females to replenish maternal reserves for future reproduction and help maximize the growth rate of neonates. This spread also indicates low synchrony of chital births and suggests that predator swamping was less important in influencing their timing of parturition. Insensitivity of the results to variable rainfall patterns lent support to the hypothesis that forage requirements determined by body mass might explain the variation in parturition strategies among large herbivores species.
Community ecology: large mammalian herbivores in a South Indian forest
There is little understanding of how large mammalian herbivores in Asia partition habitat and forage resources, and vary their diet and habitat selection seasonally in order to coexist. I studied an assemblage of four large herbivores, chital (Axis axis), sambar (Rusa unicolor), gaur (Bos gaurus) and Asian elephant (Elephas maximus), in the seasonal tropical forests of Bandipur and Mudumalai, South India, and tested predictions regarding the species’ seasonal diet browse : graze ratios, habitat selection and habitat-niche preference and overlap. Field data collected for the study included the seasonal variation in grass quality, the seasonal variation in δ13C in the species’ faeces and the seasonal variation in the species’ habitat selection and overlap using a grid-based survey. Results showed that the chital was more of a grazer in the wet season, but that it increased the proportion of browse in its diet in the dry season; the gaur was a grazer for most of the year; the sambar preferred to browse throughout the year; and that the elephant was a mixed feeder. Elephant habitat-niche breadth was high and overlapped equally with that of the other species in both seasons (Fig. below). The gaur had the most restricted habitat-niche breadth in both seasons, and it switched from the moist deciduous habitat in the dry season to the dry deciduous habitat in the wet season (Fig. below). These results offer insights into the seasonal variation in browse:graze diet ratios and the habitat-niche overlap amongst the common large mammalian herbivore species found in South India.
There is little understanding of how large mammalian herbivores in Asia partition habitat and forage resources, and vary their diet and habitat selection seasonally in order to coexist. I studied an assemblage of four large herbivores, chital (Axis axis), sambar (Rusa unicolor), gaur (Bos gaurus) and Asian elephant (Elephas maximus), in the seasonal tropical forests of Bandipur and Mudumalai, South India, and tested predictions regarding the species’ seasonal diet browse : graze ratios, habitat selection and habitat-niche preference and overlap. Field data collected for the study included the seasonal variation in grass quality, the seasonal variation in δ13C in the species’ faeces and the seasonal variation in the species’ habitat selection and overlap using a grid-based survey. Results showed that the chital was more of a grazer in the wet season, but that it increased the proportion of browse in its diet in the dry season; the gaur was a grazer for most of the year; the sambar preferred to browse throughout the year; and that the elephant was a mixed feeder. Elephant habitat-niche breadth was high and overlapped equally with that of the other species in both seasons (Fig. below). The gaur had the most restricted habitat-niche breadth in both seasons, and it switched from the moist deciduous habitat in the dry season to the dry deciduous habitat in the wet season (Fig. below). These results offer insights into the seasonal variation in browse:graze diet ratios and the habitat-niche overlap amongst the common large mammalian herbivore species found in South India.
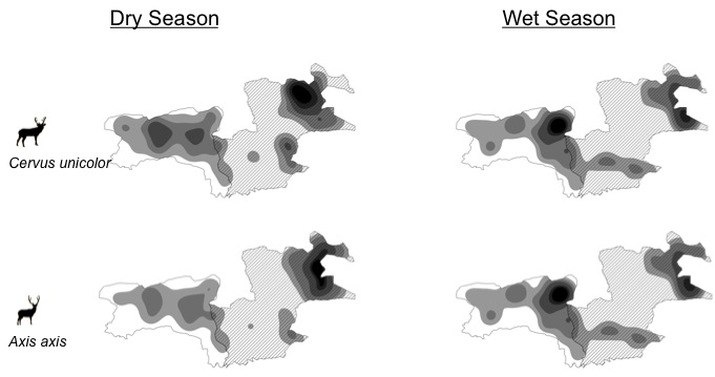
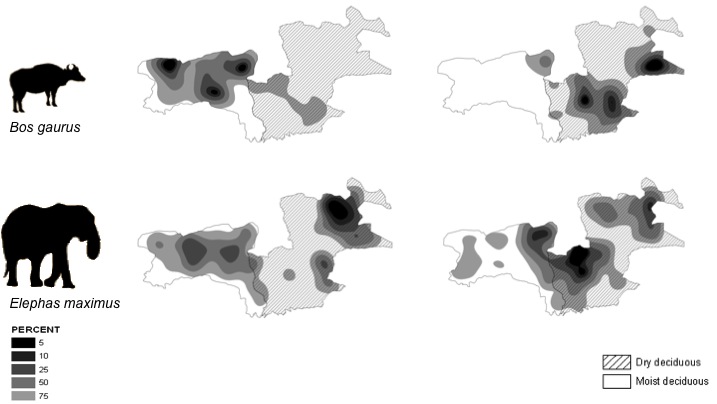
( Darker shading indicates greater presence of the species )
Bayesian state-space modeling of population time-series: the importance of observation versus process error in global ungulate populations
Population abundance data vary widely in quality and are rarely accurate. The two main components of error in such data are observation and process error. I used Bayesian state space models to estimate the observation and process error in time-series of 55 globally distributed populations of two species, Cervus elaphus (elk/red deer) and Rangifer tarandus (caribou/reindeer). I examined variation among populations and species in the magnitude of estimates of error components and density dependence using generalized linear models. Process error exceeded observation error in 75% of all populations, and on average, both components of error were greater in Rangifer than in Cervus populations. Observation error differed significantly across the different observation methods, and predation and time-series length differentially affected the error components. Comparing the Bayesian model results to traditional models that do not separate error components revealed the potential for misleading inferences about sources of variation in population dynamics.
Population abundance data vary widely in quality and are rarely accurate. The two main components of error in such data are observation and process error. I used Bayesian state space models to estimate the observation and process error in time-series of 55 globally distributed populations of two species, Cervus elaphus (elk/red deer) and Rangifer tarandus (caribou/reindeer). I examined variation among populations and species in the magnitude of estimates of error components and density dependence using generalized linear models. Process error exceeded observation error in 75% of all populations, and on average, both components of error were greater in Rangifer than in Cervus populations. Observation error differed significantly across the different observation methods, and predation and time-series length differentially affected the error components. Comparing the Bayesian model results to traditional models that do not separate error components revealed the potential for misleading inferences about sources of variation in population dynamics.
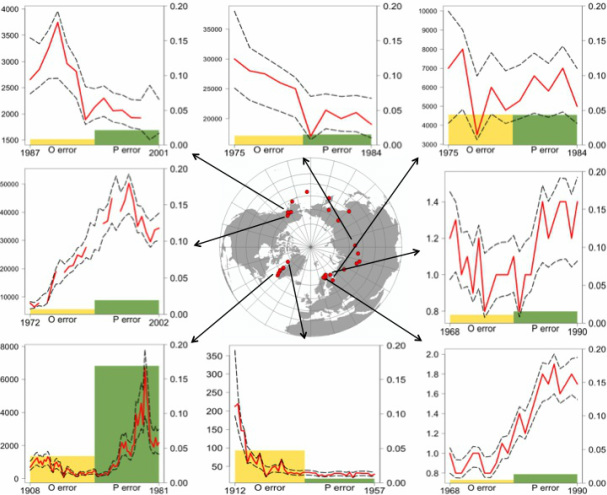
Global reindeer/caribou populations. In each graph: red line is time-series of population counts; black dashed lines are 95% confidence interval of Bayesian estimates of population time-series; yellow and green bars are Bayesian estimates of observation and process error in each population time-series, respectively.
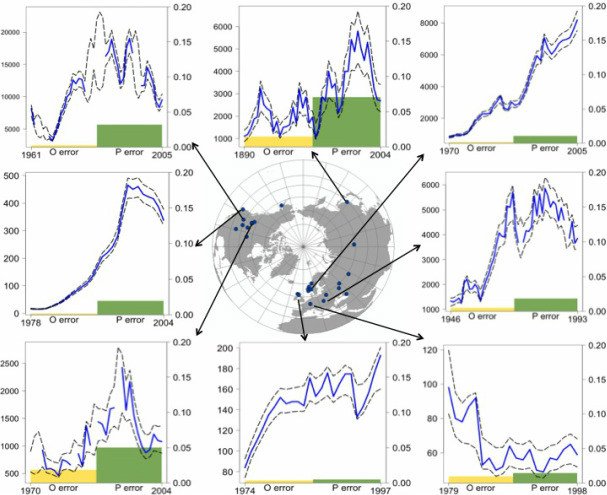
Global elk/red deer populations. In each graph: blue line is time-series of population counts; black dashed lines are 95% confidence interval of Bayesian estimates of population time-series; yellow and green bars are Bayesian estimates of observation and process error in each population time-series, respectively.
